哈尔滨河流廊道草本自生植物多样性与分布特征:低维护景观设计策略
摘要
【目的】河流廊道是城市自生植物重要的栖息地,揭示其自生植物多样性分布格局及成因,有助于完善城市植被研究体系、指导城市生物多样性保育以及低维护与可持续绿地景观营造。
【方法】采取均匀取样法和典型取样法,选取哈尔滨市何家沟和马家沟两条河流廊道的中心城区段共951个样方展开草本层自生植物调查,并解析3个大类生境和11类微生境中的自生植物多样性及分布特征。
【结果】1)共记录草本层自生植物151种,隶属于36科、112属;2)中低等城市化水平(Ⅰ~Ⅲ)下的草本层自生植物及乡土植物的\alpha多样性显著高于其他城市化水平(Ⅳ~Ⅴ)下的\alpha多样性;不同类型生境自生植物的多样性特征差异显著,入侵植物呈现均质化分布格局;3)不同生境类型所处的坡向、距水体的距离、植被结构、管理强度等因素对自生植物的生活型、发生方式以及扩散方式均产生显著影响。
【结论】为哈尔滨城市河流廊道生物多样性的保育以及低碳、低维护植物景观设计提供了新的视角,通过合理留存和利用自生植物,能最大限度地发挥河流廊道绿地的各项生态服务功能。
关键词:自生植物;生物多样性;河流廊道;城市化水平;生境异质性;哈尔滨。
1 研究方法
1.1 研究区域概况
哈尔滨市是中国最北、纬度最高的省会城市,拥有丰富的河流水系资源。何家沟全长 32.59 km,流经平房、香坊、南岗、道里4 个行政区;马家沟全长 44.3 km,流经平房、南岗、道外 3 个行政区。随着城市的不断扩张和发展,何家沟和马家沟河岸带人地关系紧张,生境破碎化严重、景观异质性持续增强,对其生物多样性的组成、结构和动态产生了重要影响。
1.2 研究方法
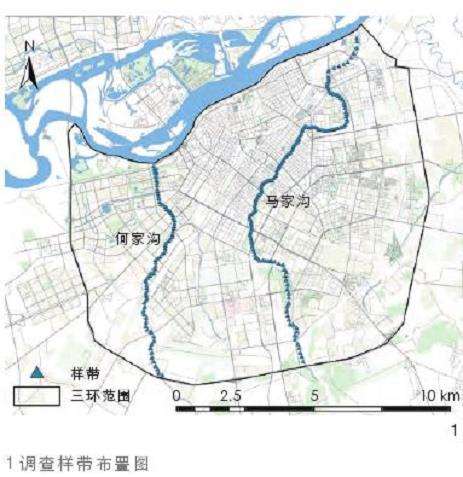
1)调研样段选取。结合两条河流的现状以及既有研究成果,选取何家沟与马家沟自松花江入江口至哈尔滨三环河段(长约 34.57 km),对于其中有连续植被覆盖的河道,以植被覆盖范围作为研究区域,对于其中无连续植被覆盖的河道,以土地利用类型明显变化边界以内的范围作为研究区域[8, 11],调查时间为2022 年 6—9 月(图 1)。
2)样方设置。以均匀取样为主,以确保样方能全面反映样地的总体基本情况。在研究区域内每间隔 200 m 做一条垂直于河岸线的样带,每条样带上每间隔 5 m 取 1m×1m 的样方记录植物。样带之间出现新的物种和新生境时,也设置典型样方进行记录。本次调查共设置草本群落样方 951 个,记录所有样方内植物的物种名、株数、高度、盖度、是否自生。本研究中自生植物包括城市杂草、栽培植物中的木本植物更新幼苗[14],以及废置地、硬质铺装和护岸缝隙等生境内生长的木本和草本植物;对于胸径超过 2 cm 的木本植物、有明显人工栽培管理痕迹的草本植物,均认定为栽培植物。
参考前人对植被分布驱动因素的研究[4, 6],同时记录样方距河岸线的水平距离,样方所处生境的类型、坡度、坡向、宽度、管理强度以及内部植被结构、木本植物盖度。将坡向 22.5°~67.5°记为北,67.5°~112.5°记为东北,每 45°依次记录为东、东南、南、西南、西、西北。管理强度按照种植、灌溉、收割和基础设施建设等人工管理行为强度划分为等级 1~5 。植被结构划分为草地、灌-草、乔-草、乔-灌-草。
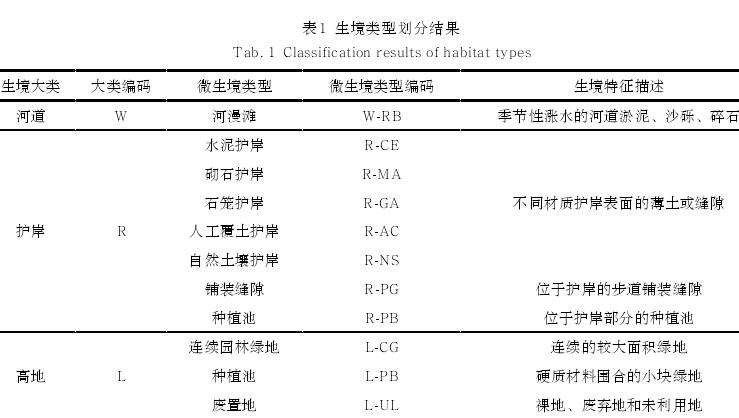
3)生境类型划分。所有样方内自生植物生境按照所处河道的位置划分为河道、护岸和高地三大类[15],并在此基础上按照生境本底特征及使用功能进一步划分为 11 类微生境(表 1)。
4)城市化水平指数计算。运用 Arc GIS 10.8软件中的最大似然法,将 2020 年分辨率 30 m的 Landsat 系列卫星影像解译为城市建设用地、水体、植被、耕地和裸地 5 个土地覆盖类型。以每条样带中点为中心、500 m 为半径,计算样带周边不透水率(即建设用地面积占比),并将样带周边不透水率按照 0.2 的数值间隔将0~1.0 划分为Ⅰ~Ⅴ5 个等级,由低到高代表该样带所处位置的城市化水平。
1.3 数据处理
1)采用重要值(P)作为评价各物种优势程度的指标,计算式为式中:hi 为物种 i 的相对高度,ci 为物种 i 的相对盖度。
2)\alpha 多样性是指某个群落或生境内部的物种多样性,选取 \alpha 多样性指数中的 Patrick丰富度指数(R)、Shinnon-Wiener 多样性指数(H)和 Pielou 均匀度指数(J)作为测度指标。其中 Patrick 丰富度指数是指一个群落中所有实际物种的数目(S);Shinnon-Wiener多样性指数来源于信息理论,其值愈大,群落所含的信息量愈大,计算式为Pielou 均匀度指数是群落中不同物种的多度 (生物量、盖度或者其他指标)分布的均匀程度,计算式为J = H/ln(Pi) , (3)式(2)、(3)中,Pi 为物种 i 的重要值。
\alpha 多样性指数利用 R 4.3.1 软件中的 Vegan包进行计算。不同城市化水平下与不同生境间 \alpha 多样性的差异比较以及不同生境的环境因素中分类变量对比均采用 Kruskal-Wallis 差异性检验,数值变量对比均采用 Spearman 相关性检验,检验过程利用 R 4.3.1 软件中的 Rstatix包计算,检验结果利用 plot 包与 Pheatmap 包进行可视化。自生植物的物种来源、生活型、发生方式、扩散方式、是否为乡土植物、入侵植物等信息主要参考《中国植物志》《黑龙江省植物志》《中国入侵植物名录》《中国杂草原色图鉴》《黑龙江省主要乡土树种名录》《黑龙江省主要乡土草种名录》、中国植物物种信息数据库及 SID 数据库进行确定,入侵植物参考中国外来入侵物种信息系统(Invasive Alien Species of China, IASC)划分为恶性入侵、严重入侵、局部入侵类、一般入侵和有待观察 5 类。
2 结果与分析
2.1 草本层自生植物的物种组成
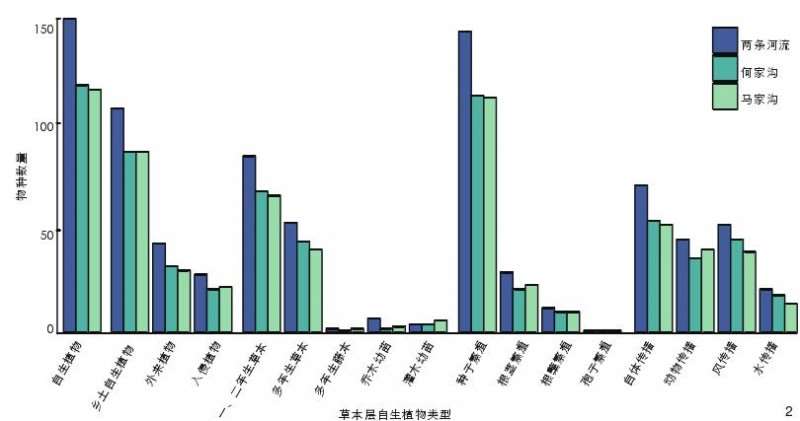
本次共记录草本层自生植物 151 种,隶属于 36 科、112 属,不同河流廊道中草本层自生植物的组成特征存在一定差异(图 2)。
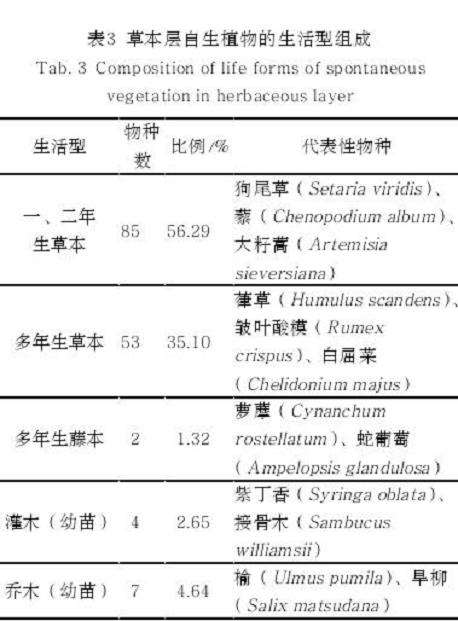
从科属构成上看,菊科(Asteraceae)物种数量最丰富,其次是禾本科(Gramineae)和蓼科(Polygonaceae),是两条河流廊道中草本层自生植物的优势科(表 2);从生活型来看,一、二年生草本自生植物占主导地位,其次为多年生草本(表 3);物种来源以乡土自生植物为主,入侵自生植物中恶性入侵类有4 种,包括小蓬草(Erigeron canadensis)、鬼针 草( Bidens pilosa)、 反 枝 苋 ( Amaranthusretroflexus)、 圆 叶 牵 牛 ( Ipomoea purpurea,表 4);不同繁殖方式中,种子繁殖占绝对优势,孢子繁殖的物种仅问荆(Equisetum arvense)1 种(表 5);从扩散型来看,自体传播的物种最多,风传播和动物传播自生植物物种数相近(表 6)。

2.2 不同城市化水平草本层自生植物多样性特征
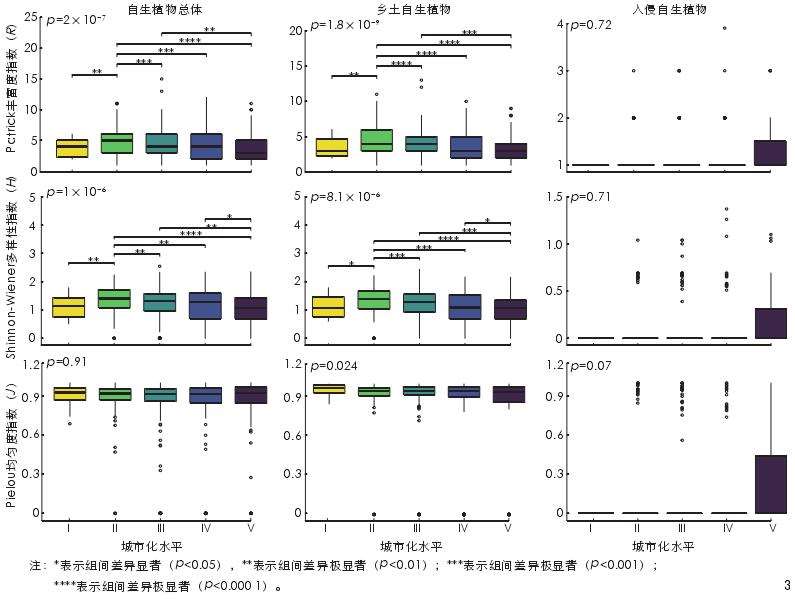
草本层自生植物的 Patrick 丰富度指数 (自生-R)和 Shinnon-Wiener 多样性指数(自生-H)均值在城市化水平Ⅱ中最高,且在不同城市化水平上的差异显著(p<0.05)。乡土自生植物的 Patrick 丰富度指数(乡土- R)和 Shinnon-Wiener 多样性指数(乡土-H)的最高均值同样出现在城市化水平Ⅱ中,且乡土-R、乡土-H 和 Pielou 均匀度指数(乡土-J)在不同城市化水平上的差异均显著(p<0.05)。入侵植物的 Patrick 丰富度指数(入侵-R)和Shinnon-Wiener 多 样 性 指 数 ( 入 侵 -H) 和Pielou 均匀度指数(入侵-J)在不同城市化水平上的差异均不显著(p≥0.05,图 3)。
2.3 不同生境类型草本自生植物多样性特征
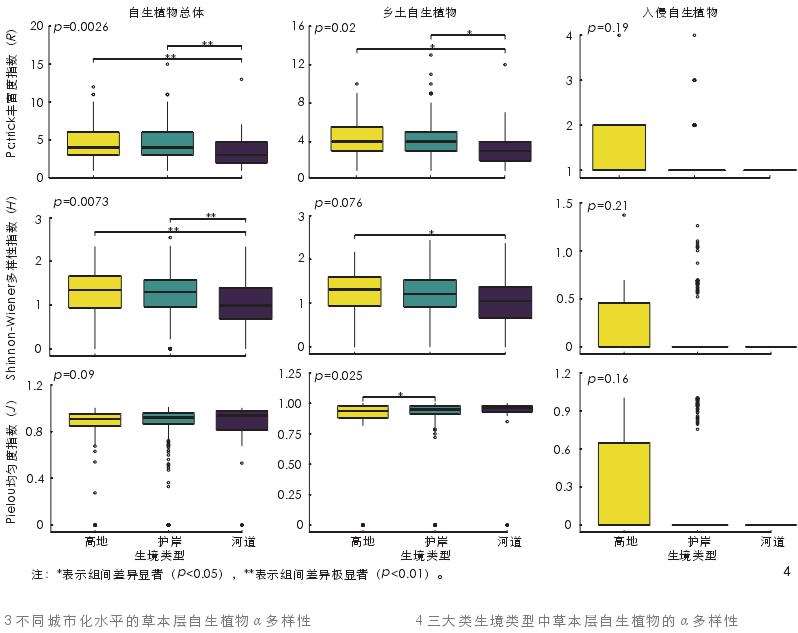
自生-R、自生-H、乡土-R 和乡土-H 在三大类生境类型中的差异性显著(p<0.05),且其均值排序均为高地>护岸>河道;自生-J、乡土-J、入侵-R、入侵-H 和入侵-J 在三大类生境类型中均无显著差异(p≥0.05,图 4)。
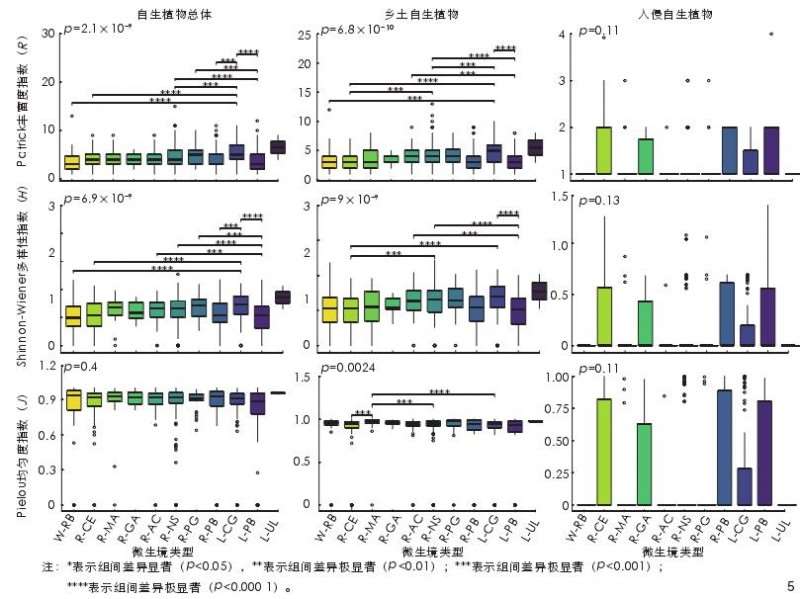
不同微生境类型中自生-R、自生-H、乡土-R 和乡土-H 的差异性显著(p<0.05),其中高地废置地、高地连续园林绿地、护岸铺装缝隙生境的 \alpha 多样性指数均值最高。乡土-J在不同微生境类型间的差异性显著(p<0.05),而入侵-R、入侵-H 和入侵-J 在不同微生境类型中的差异性均不显著(p≥0.05,图 5)。
2.4 自生植物多样性分布格局的驱动因素
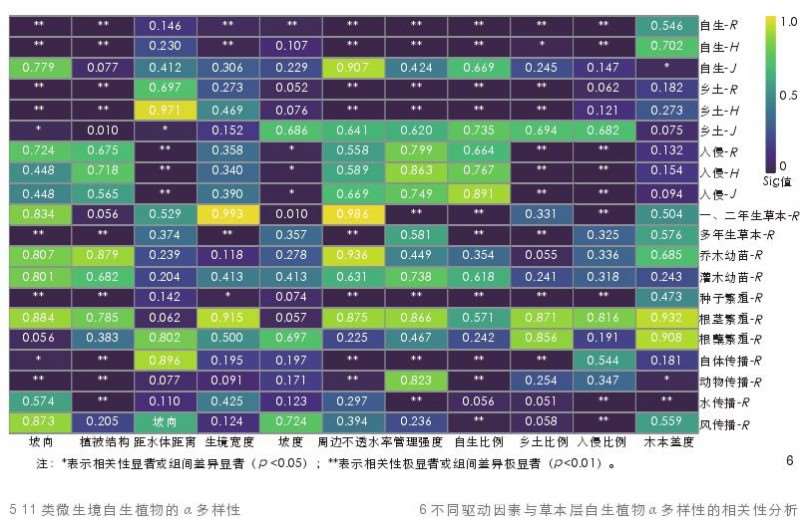
对不同驱动因素中数值变量与草本层自生植物多样性进行 Spearman 相关性检验,发现自生-R 与自生-H 与周边不透水率、管理强度均显著负相关(图 6)。从物种来源来看,入侵自生植物的 \alpha 多样性指数均与样方距水体的距离显著正相关,与乡土物种比例显著负相关;从生活型来看,一、二年生草本-R与管理强度显著负相关;从发生方式来看,种子繁殖-R 与自生比例和入侵比例显著正相关;从扩散方式来看,自体传播-R 与周边不透水率和管理强度显著负相关,动物传播-R在西南坡向显著高于东坡向和北坡向,水传播-R 与入侵比例显著正相关,与管理强度和木本植物的盖度显著负相关,风传播-R 与入侵比例显著正相关。
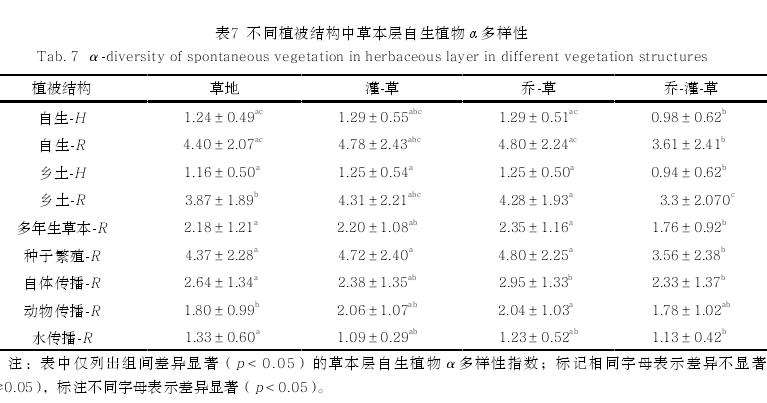
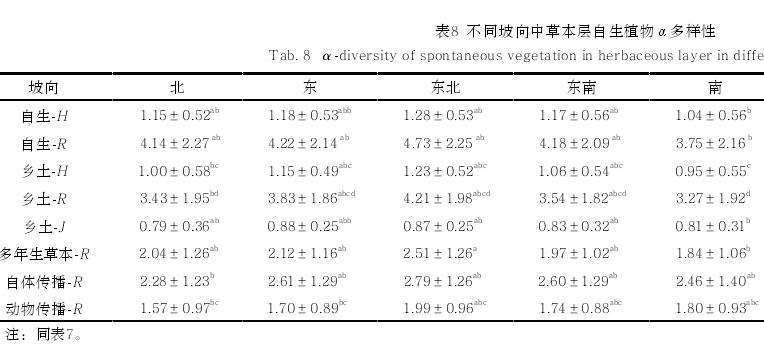
对不同驱动因素中类别变量与草本层自生植物 \alpha 多样性指数进行Kruskal-Wallis H 检验,发现多数类别草本层自生植物 \alpha 多样性在乔-草和草地植被结构中高于在乔-灌-草植被结构中(表 7),多数类别草本层自生植物 \alpha 多样性在南坡向最低 (表 8)。
3 讨论
3.1 草本自生植物的物种组成特征
本研究在哈尔滨市何家沟、马家沟河流廊道中心城区段共记录草本层自生植物 151 种,约占哈尔滨三环内城市绿地维管束植物种类数(407 种)[16] 的 37%。由此可见,河流廊道自生植物是城市生物多样性的重要组成部分。研究区域共记录乡土自生植物 108 种,占自生植物物种总数的 71.52%,和其他类似研究结果[8, 11] 接近,表明河流廊道对乡土植物的保育具有积极的意义。草本层自生植物的生活型主要以一、二年生草本为主,共 85 种,它们能通过在短时间内完成生活史,以适应河流廊道水湿条件变化较大且易受人类干扰的生态环境。
草本层自生植物的扩散方式以自体传播为主,其次为风传播、动物传播和水传播,与南京市公园绿地的研究结果一致[17]。由于城市建筑的形态会影响城市风环境,不同的风向和风速对植物种子的传播距离存在影响[18],同时可以传播种子的城市动物相对较少[19],所以城市河流廊道中风传播和动物传播自生植物的多样性会低于原始自然生态环境区域。
3.2 自生植物多样性对异质性生境的响应与适应
研究结果表明,自生植物是研究城市化与城市生态系统互馈机制的绝佳对象。哈尔滨河流廊道草本层自生植物的 \alpha 多样性指数在不同城市化水平下差异显著,在中低等城市化水平(Ⅰ~Ⅲ)的区域达到最高,而入侵自生植物的 \alpha 多样性指数在不同城市化水平上无显著差异,与北京温榆河入侵植物的分布规律相似[11]。这表明乡土自生植物对城市当地的生物多样性具有支持作用[20],而入侵物种因具有更强的扩散能力和适应能力,加上人类干扰活动而加速进入河流廊道,同时河流廊道也促进了入侵植物的扩散过程,进而形成了入侵物种同质化的分布格局[21]。
城市河流廊道是水陆系统的交错带,生境类型丰富,异质性生境的形成对自生植物多样性分布格局的影响较为显著。河漫滩生境是水生及湿生植物重要生境类型,但该生境易受雨洪干扰和河道清淤管理的影响,导致其草本层自生植物多样性最低;高地生境类型中废置地和连续园林绿地这两类面积较大的微生境维持着最高的草本层自生植物 \alpha多样性,符合生态学中的“种-面积”关系规律。
草本层自生植物 \alpha 多样性在高地各类微生境和护岸中的自然土壤护岸、人工覆土护岸、种植池这些以土壤为基底的微生境中高于在砌石护岸、石笼护岸、水泥护岸这些硬质护岸生境中,是因为河岸的硬化工程不仅破坏了沿岸动植物的天然栖息地,同时也隔断了沿岸的雨水与河水的自然循环,对河流生态系统及生物多样性造成了不可逆转的影响[22]。
自生植物 \alpha 多样性同时也受到不同生境类型中生物因素和非生物因素的影响。从不同植被结构来看,草地中的草本层自生植物\alpha 多样性指数最高,与前人研究结果[23] 一致。从不同坡向可以看出,西南坡向的 \alpha 多样性指数水平最高,是由于西南坡属于阳坡和半阳坡生境,阳光充足,利于草本层中阳生植物发育[24]。
此外,越靠近河流廊道边缘的生境类型,入侵植物的 \alpha 多样性指数越高,有学者在青岛唐岛湾的研究中同样发现道路边缘绿地入侵植物物种数量高于其他位置[25],因此河流廊道边缘应该是入侵植物预防监控的重点区域。然而,生境内部及周边不透水率与入侵植物 \alpha 多样性的相关性并不显著,可能与入侵植物的 \alpha 多样性对 500 m 半径尺度下的不透水率不敏感有关,有待从更多尺度上开展进一步研究。一、二年生草本,种子繁殖和水传播植物受管理强度的影响显著,其原因在于一、二年生草本植物生命周期短,对人类干扰活动的响应更迅速;种子繁殖植物易受除草时间和频率的影响;水传播植物在研究区域内多生长于河漫滩和水岸,更容易受河道清淤作业的影响。
3.3 生物多样性提升与低维护植物景观营造的策略
基于自生植物在不同城市化水平、生境类型以及生态环境特征下呈现出的分布规律和植物景观特征,本研究分别从生境营造和物种选择两个方面提出城市河流廊道生物多样性提升和低维护植物景观营造的策略,有助于在城市景观建设过程深入贯彻低维护、可持续植物景观设计理念,真正发挥城市自生植物在绿色低碳与可持续发展中的关键作用。
3.3.1 生境营造策略
1)在满足城市河道防洪功能和安全功能的前提下,降低部分河段的沟渠化程度。在河流入江口周边河段和城市居民生产生活频繁的河段,应以防洪功能和安全功能为主。在承担次要泄洪功能的河段,以抛石、石笼、混凝土球、空心砌砖等生态护岸代替混凝土、浆砌石块等传统硬质护岸,同时可以适当扩宽河道,增加横截面以提升蓄洪量[26]。
在上游低城市化水平河段,应尽可能保护河流廊道的自然形态和原有土壤底质,从而形成河漫滩、河岸、护坡、高地的多级河道结构,营造丰富多样的微生境格局和近自然河流景观,还有利于降低流速、延长滞留时间、增加河道容量和生物多样性。
2)保护生境的连续性与完整性。不同生境类型自生植物多样性与单个生境的面积大小和完整性紧密相关[27],对废置地、连续园林绿地和其他面积较大的生境斑块应予以优先保护。对于被道路和其他硬质铺装分割的生境类型,建议采用嵌草铺装或步石打破硬质界面,以提升各类生境间的连通度,增强物种迁移能力。
3)提升生境内植被结构的多样性。在种植池、连续园林绿地等以人工栽培植被为主的微生境中,增加乔-草和草地等多种植被结构能够有效降低郁闭度,从而提升草本植物的多样性。与此同时,为给鸟类、蝶类、蜂类等小型动物提供活动、觅食、隐蔽和栖息的条件[28],在所有生境类型中都应保留足够面积的乔-灌-草和灌-草结构[29]。
4)保护部分生境的独特性。研究结果显示,西南坡向的生境类型往往有着较高的物种多样性水平,例如在河道景观建设过程中在西南坡向可以采用低维护的自生植物;铺装缝隙生境类型的土壤贫瘠且易受到踩踏,但对其中生存的抗逆性较强的物种建议适当保留,在提高城市生物多样性的同时,也有助于形成富有自然野态的植物景观。
3.3.2 物种选择策略
1)选取乡土植物和低入侵风险外来植物。对于恶性入侵类、严重入侵类和局部入侵类,应定期调查、监测,采取物理防治的同时使用乡土植物对该类入侵物种进行生态替代,以河流廊道边缘为重点调查监测区域。而对于有观赏价值的一般入侵和有待观察类入侵物种,如草木樨(Melilotus suaveolens)、苜蓿虎尾草、毛酸浆(Physalis philadelphica)、菊芋(Helianthus tuberosus)、香附子(Cyperusrotundus)、荠(Capsella bursa-pastoris)、鹅肠菜 ( Stellaria aquatica)、 地 锦 ( Parthenocissustricuspidata),可与竞争力较强的乡土自生植物共同作为植物群落构建材料。
2)选取生境适应性强的自生植物。一般来说,城市化水平较高(Ⅳ~Ⅴ)的河段受人类活动干扰较大、硬质化生境较多,适合使用一、二年生,种子繁殖或自体传播的自生草本植物作为植物景观营造的材料。对于不同生境类型来说,滨水生境适合使用水传播、耐水湿的自生植物材料,而远离水岸的生境则推荐使用其他扩散方式的物种;硬质生境适合使用抗逆性较强、植株较小、生命周期较短的自生植物材料;以土壤为底质的生境对植物的发生方式和传播方式的选择则相对宽泛(表 9)。
对于不同植被结构的微生境来说,在草本层生境中推荐采用自体传播和风传播的自生植物种类,在密度较大的灌木层或乔木层生境建议减少风传播的植物比例,增加自体传播和动物传播且较为耐阴的自生植物物种比例。在东南、南、西南、西、西北坡向中使用喜阳的自生植物,在东北、东、北坡向中使用喜阴、耐阴的自生植物。
4 结论
城市河流廊道是自生植物重要的栖息地,自生植物不仅是城市生物多样性的重要组成部分,也是研究城市化与城市生态系统互馈机制的绝佳对象。本研究以哈尔滨市两条河流廊道的草本层自生植物为研究对象,探索了自生植物的 \alpha 多样性分布特征及其成因。研究结果显示,哈尔滨城市河流廊道的草本层自生植物资源丰富,在不同城市化水平上,草本层自生植物的 \alpha 多样性差异显著,且生境类型、坡向、植被结构类型等是影响其 \alpha多样性分布的关键因子。
乡土自生植物对维持河流廊道生物多样性具有积极作用。在未来低碳可持续发展的背景下,在进行城市河流廊道景观建设与管理过程中,应针对自生植物的适应性策略开展生境营造和植物群落设计,结合适度的管理养护以及对外来入侵植物的密切监测,更好地发挥河流廊道对城市生物多样性的支持功能。在未来的研究中,可通过多季节调查和群落调查,进一步探索城市河流廊道草本层自生植物的时空分布特征。


